![]()
Aroids and other genera in the Collection
Take the Tour Now?
Orchids
The
Exotic Rainforest
Plants in
the Exotic Rainforest Collection
The images on this website are copyright protected. Please contact us before any reuse.
Detailed information on Growing Anthurium Species
Click this Link
The
Exotic Rainforest is a private botanical garden.
Within our collection we have many species of Anthurium. If you are seeking other photos, click this link:
New: Understanding, pronouncing and using Botanical terminology, a Glossary
Anthurium sagittatum (Sims) G. Don

Anthurium sagittatum (Sims) G. Don
Synonyms: Anthurium
rubrinervium, Anthurium aeranthe, Anthurium alienatum,
Anthurium amazonicum,
Anthurium cordatosagittatum, Anthurium humboldtianum subsp viridispadix,
Anthurium humboldtianum,
Anthurium polyrrhizon, Anthurium sororium,
Anthurium terrestre,
Pothos rubrinervius, Pothos sagittatus, Pothos suaveolens
The scientific information on
this species was primarily extracted from the published field notes of
aroid botanist
Dr. Thomas B. Croat Ph.D., P.A. Schulze Curator of Botany at the Missouri
Botanical Garden in St. Louis.
Additional material was extracted from the paper A Comparative Analysis of the Diversity,
Distribution,
and Biogeography of Araceae in Southeastern Peru by Jorge Linga
Chavez
Anthurium sagittatum
is currently listed on the RED LIST of Endangered Species.
Published to science in 1839 by Scottish botanist George Don (1798 to 1856) in Hortus Britannicus
#633, Anthurium
sagittatum
is placed in Anthurium section Cardiolonchium. Section
Cardiolonchium is characterized by a velutinous (velvety) sheen to the
leaf blades. The scientific term
velutinous
 indicates the leaf surface is covered with a short fine and densely
silky pubescence (hair) which causes the blade to appear to be velvety.
However, due to natural variation this characteristic is not always apparent in
Anthurium sagittatum. Botanically the species is considered
sub-velvety and the species is often known to collectors as Anthurium
rubrinervium
(correctly a synonym).
indicates the leaf surface is covered with a short fine and densely
silky pubescence (hair) which causes the blade to appear to be velvety.
However, due to natural variation this characteristic is not always apparent in
Anthurium sagittatum. Botanically the species is considered
sub-velvety and the species is often known to collectors as Anthurium
rubrinervium
(correctly a synonym).
Anthurium sagittatum is found largely in most of Colombia and Ecuador but also grows in the rain forests of Peru ranging from the Amazonas region to Loreto and Madre de Dios as well as in western and northern Brazil, Suriname, Guiana, French Guiana and almost certainly Venezuela. Anthurium sagittatum has been found in nature at elevations ranging from 110 to as high as 3100 meters (330 to over 10,000 feet) above sea level but is observed more frequently at lower elevations ranging from 100 to 1500 meters (325 to 5,000 feet). This Anthurium species is not known to grow epiphytically on the branches of trees but instead often grows in groups as a terrestrial species in both primary and virgin rain forests.
Anthurium sagittatum is frequently found along trails, roads, rocky cliff banks and especially along the banks of streams and rivers. The species appears to prefer somewhat shadowy wet places in subtropical forests and is sometimes found in depressions and swampy areas. In Ecuador Anthurium sagittatum is reported to be commonly observed in three fourths to full shade but never in full sun.
The name "sagittatum" originates from the botanical term sagittate which indicates a leaf that is "arrow shaped". It is possible the species grows as a hemiepiphyte (hem-a-EPA-fit) (a species that begins life in the soil and then climbs a tree) but scientific documentation cannot be located by this author to support that possibility. The photo to the left of Anthurium sagittatum by Elizabeth Campbell was taken near Mera, Ecuador traveling towards Llanganates Bajo in Pastaza province. The specimen is approximately 1100 meters (3,600 feet) above sea level and is fully terrestrial growing beside a gravel road in a drainage ditch of standing water.
 Aroid including Anthurium species are known to be extremely variable
and not every specimen
of Anthurium sagittatum will always appear exactly the same.
Variations may be found in the general shape and texture of the leaves as well
as other
characteristics of the plant including the coloration of the venation. A link to an
explanation of the natural variations in aroids can be found at the bottom of
this article.
Aroid including Anthurium species are known to be extremely variable
and not every specimen
of Anthurium sagittatum will always appear exactly the same.
Variations may be found in the general shape and texture of the leaves as well
as other
characteristics of the plant including the coloration of the venation. A link to an
explanation of the natural variations in aroids can be found at the bottom of
this article.
Since early botanists did not have access to the scientific records and computers we have available today they sometimes granted multiple new names to the same species. All the names listed as synonyms at the top of this page have been published as a unique scientific species but all have now been sunk into synonymy under the originally published name Anthurium sagittatum. A synonym is simply the same species published with a new name after the original scientific name was published. In botany the first published name becomes the accepted name and all other published names for that same plant become synonyms. Science does not simply change names as is sometimes believed by plant collectors but instead relies on the rules of botany to grant the first properly published name to be the accepted scientific name.
Based on the Linnean rules of nomenclature established by Swedish physician, zoologist and botanist Carl Linnaeus (1707–1778) the scheme of binomial nomenclature (two names composed of the correct genus followed by a species name) has a precise set of rules that must be followed in order to determine the proper name for any species. As can be observed, some of the names originally used were published with the wrong genus. The system Linnaeus devised defines the kingdom, phylum, class, genus, and species of any living organism. One of the most basic rules of taxonomy is the earliest published scientific name correct to genus becomes the accepted name. In this case the accepted species name is Anthurium sagittatum which was published in 1839.
 My friend naturalist Joep
Moonen (pronounced Yupe) who lives and works in French Guiana in the northeastern
corner of South America offered this information
based on his field studies of the growth of Anthurium sagittatum.
Joep has known the plant as Anthurium
rubrinervium,
"It is the most
common
of the two terrestrial Anthurium species of French Guiana, the other being
Anthurium thrinax. Your Anthurium is not common and grows in groups in
very humid circumstances preferably on rocks near criques in the shade.
In the Kaw Mountains in northeast French Guiana it grows on hill slopes in primary forest. In French
Guiana it is
extremely humid with an annual rainfall in the Kaw Mountains of about 20 feet (6 meters)!"
Joep guides botanists,
taxonomists, zoologists and visitors into the
rain forests of French Guiana to study the natural flora and fauna. He has
personally found a number of plants not previously known to science and has several aroid species named in his
honor including both Anthurium moonenii and Philodendron moonenii.
My friend naturalist Joep
Moonen (pronounced Yupe) who lives and works in French Guiana in the northeastern
corner of South America offered this information
based on his field studies of the growth of Anthurium sagittatum.
Joep has known the plant as Anthurium
rubrinervium,
"It is the most
common
of the two terrestrial Anthurium species of French Guiana, the other being
Anthurium thrinax. Your Anthurium is not common and grows in groups in
very humid circumstances preferably on rocks near criques in the shade.
In the Kaw Mountains in northeast French Guiana it grows on hill slopes in primary forest. In French
Guiana it is
extremely humid with an annual rainfall in the Kaw Mountains of about 20 feet (6 meters)!"
Joep guides botanists,
taxonomists, zoologists and visitors into the
rain forests of French Guiana to study the natural flora and fauna. He has
personally found a number of plants not previously known to science and has several aroid species named in his
honor including both Anthurium moonenii and Philodendron moonenii.
When attempting to identify any
aroid species the specific characteristics of the species being researched are
important. This article attempts to explain and illustrate these
characteristics in a way any collector can use to verify whether or not a plant
is or is not a specific species.
 The leaf blades of Anthurium
sagittatum are dark green and semi-glossy to weakly glossy on the
upper (adaxial) surface but substantially paler and matte on the
underside (abaxial surface). The leaves stand erect. The upper
surface may be a mixture of velvety
and matte (sub-velvety) and are subcoriaceous as well as
somewhat papyraceous (papery). Subcoriaceous
indicates the blades are just less than leathery to the touch while
coriaceous indicates the leaves are thick.
The leaf blades of Anthurium
sagittatum are dark green and semi-glossy to weakly glossy on the
upper (adaxial) surface but substantially paler and matte on the
underside (abaxial surface). The leaves stand erect. The upper
surface may be a mixture of velvety
and matte (sub-velvety) and are subcoriaceous as well as
somewhat papyraceous (papery). Subcoriaceous
indicates the blades are just less than leathery to the touch while
coriaceous indicates the leaves are thick.
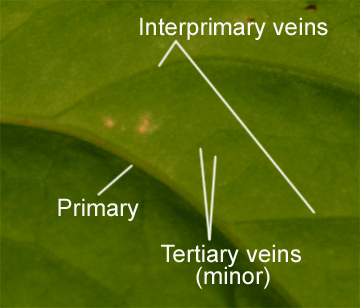
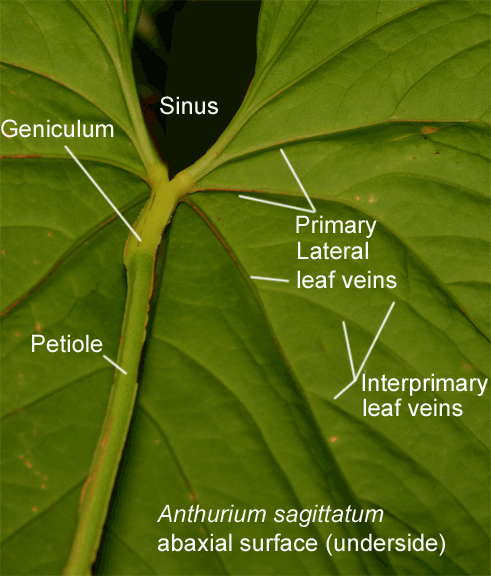 The veins on a leaf are divided
into classifications known as the basal ribs, midrib, primary lateral veins,
interprimary veins and
tertiary (or minor) veins (see both photos above right and to the left above and
below), In the case
of Anthurium sagittatum the midrib, which runs down the center of the leaf
blade, is narrowly raised and
roughly
concolorous (single colored) on the adaxial (upper) surface.
As the midrib approaches the lower blade tip the leaf rounds to a slender
point known as being acute. The primary lateral
leaf veins are slightly sunken (almost appearing to be somewhat quilted).
Although sunken into the blade's upper surface the primary veins themselves are
narrowly raised. The
veins are acutely raised on the abaxial surface (underside). Both the midrib and primary
veins may also be reddish in color on the underside of the leaf. This
reddish venation is where the synonym
Anthurium rubrinervium
(red nerve) received its name. The
tertiary (minor) veins are partially raised on the underside of the leaf and are
only slightly conspicuous but are often closer to obscure.
The veins on a leaf are divided
into classifications known as the basal ribs, midrib, primary lateral veins,
interprimary veins and
tertiary (or minor) veins (see both photos above right and to the left above and
below), In the case
of Anthurium sagittatum the midrib, which runs down the center of the leaf
blade, is narrowly raised and
roughly
concolorous (single colored) on the adaxial (upper) surface.
As the midrib approaches the lower blade tip the leaf rounds to a slender
point known as being acute. The primary lateral
leaf veins are slightly sunken (almost appearing to be somewhat quilted).
Although sunken into the blade's upper surface the primary veins themselves are
narrowly raised. The
veins are acutely raised on the abaxial surface (underside). Both the midrib and primary
veins may also be reddish in color on the underside of the leaf. This
reddish venation is where the synonym
Anthurium rubrinervium
(red nerve) received its name. The
tertiary (minor) veins are partially raised on the underside of the leaf and are
only slightly conspicuous but are often closer to obscure.
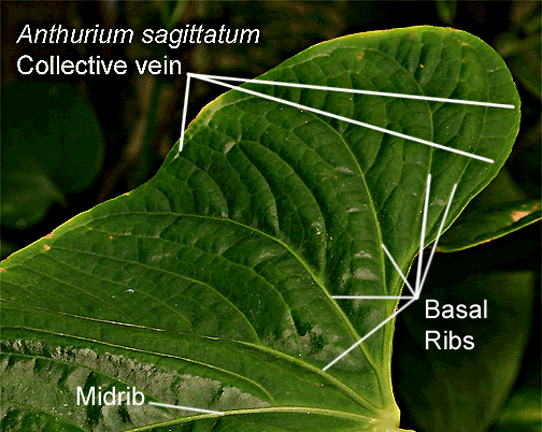
All Anthurium
species possess
a collective vein which runs near the edge of the blade.
The
 collective vein is a specialized vein to which other veins connect.
In the case of
Anthurium sagittatum the vein runs around the
entire
circumference of the species' blade. In some Anthurium species
the collective vein may
run only partially around the blade's circumference.
collective vein is a specialized vein to which other veins connect.
In the case of
Anthurium sagittatum the vein runs around the
entire
circumference of the species' blade. In some Anthurium species
the collective vein may
run only partially around the blade's circumference.
The collective vein along with the geniculum (see below) are primary characteristics used to determine if a specimen is or is not an Anthurium. Genera such as Philodendron do not possess either a collective vein nor a geniculum while other aroid genera may collect one or the other.
The stem is the part of the
plant that forms buds, petioles (thus leaves), roots, and the peduncles that support
an inflorescence (thus
flowers). The stem (photo, right) is the main axis of the plant and is the plant's base
which provides support.
Although the majority of the
stem grows above ground portions may also exist beneath the
surface. The purpose of the stem is to collect and store water and nutrients
which are absorbed by the specimen's roots. The roots grow (along with the petioles) from the nodes along
the stem's length. In many
Anthurium species those roots never touch the
soil but
dangle in the air since many species grow suspended on the trunk or limbs of
a
 tree. Anthurium sagittatum is
terrestrial and rather than growing in the forest canopy grows in the soil. Along the length of the stem
which is short there can be found nodes separated by
internodes which are also short. The internodes typically measure 1.5 to 3 cm in diameter. The
internodes are simply a stem segment between two nodes. The
roots of Anthurium sagittatum are light brown to brown in color.
tree. Anthurium sagittatum is
terrestrial and rather than growing in the forest canopy grows in the soil. Along the length of the stem
which is short there can be found nodes separated by
internodes which are also short. The internodes typically measure 1.5 to 3 cm in diameter. The
internodes are simply a stem segment between two nodes. The
roots of Anthurium sagittatum are light brown to brown in color.
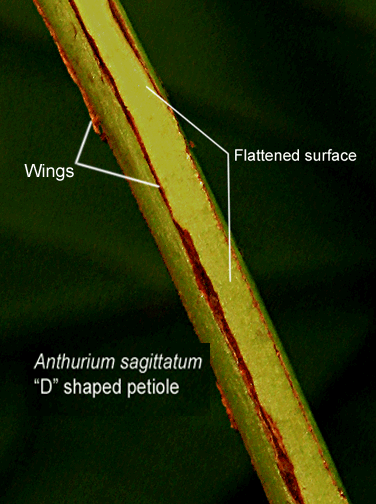
 The
stalks that support the leaves are known as petioles (detail photo, left). Despite common misconceptions the petiole is not the "stem", a
term commonly used incorrectly by plant collectors. The petiole is simply a stalk that
connects the leaf blade at the top of the petiole to the stem at its bottom. The petiole grows
from a bud on the stem and transfer water along with nutrients from the stem to the leaves. The
blades then convert carbon dioxide in the air into oxygen for other living
organisms such as humans and rain forest animals to breathe.
The
stalks that support the leaves are known as petioles (detail photo, left). Despite common misconceptions the petiole is not the "stem", a
term commonly used incorrectly by plant collectors. The petiole is simply a stalk that
connects the leaf blade at the top of the petiole to the stem at its bottom. The petiole grows
from a bud on the stem and transfer water along with nutrients from the stem to the leaves. The
blades then convert carbon dioxide in the air into oxygen for other living
organisms such as humans and rain forest animals to breathe.
The petioles of Anthurium sagittatum are tinged violet-purple and are "D" shaped plus faintly ribbed on the upper surface (adaxially). The description "D" shaped refers to the petiole when viewed as having been cut as a cross section. Once cut the upper surface can be seen as flat while the balance of the petiole is terete (round) making it appear as a capital letter D, That upper surface is both somewhat flattened and sulcate. The botanical term sulcate may indicate either a channel that runs along the upper surface of the petiole or fine parallel grooves that run along that surface. The petioles are angular winged and may possess as many as 8 wings (see photo, above right). The base of the petioles are tinged reddish brown and are sharply 3 ribbed but may also have 4 to 5 sides as they intersect the stem. The petioles may also be striate which indicates they possess parallel lines (not grooves) that run along the petiole's axis (length).
All Anthurium
species possess an organ known to botanists as a geniculum (see
photo, below left). The word originates from the scientific term geniculate
which means to be bent sharply like a knee. The geniculum is found at the petiole's apex
(top) just beneath the connection to the blade and is normally slightly larger than the
diameter of the petiole. The geniculum
often appears as if it were a
a slightly swollen "gland" and on Anthurium
sagittatum is somewhat lighter in coloration. The geniculum acts
as a knee or elbow and permits the leaf to rotate and change its orientation in
order to better
collect light. In the majority of Anthurium
species the geniculum is easily observed but in a few it is
 somewhat obscure but
almost always can be detected by feel.
somewhat obscure but
almost always can be detected by feel.
The cataphylls are bract-like modified leaves that surround any
newly emerging leaf blade and its petiole. The purpose of
the cataphyll is primarily to protect the new leaf
as it develops, The cataphylls of Anthurium sagittatum are
bluntly ribbed once they emerge and
are semi persistent as well as split in half once they age. The cataphylls
of Anthurium sagittatum are
are triangular lanceolate in shape.
Triangular lanceolate indicates the cataphylls are both triangular and lance
shaped as in the shape of a knife. They then age to a
tannish brown coloration (see photo, right) and persist at the base of the
petiole for a period of time before finally becoming deciduous and dropping
from the plant.
Anthurium
species are members of the larger
plant family known as Araceae. Members of the genera of Araceae are
also commonly known as aroids. An aroid is a
plant that reproduces by producing an inflorescence which consists
of a spathe and spadix along with other near microscopic parts. The inflorescence is
supported by a stalk known as the
peduncle which is terete (round) and is either shorter or
approximately as long as
a petiole,
Many people believe the spathe is a "flower" which is incorrect. The spathe is a modified leaf whose purpose is to protect the spadix at the center of the inflorescence during development. When sexually mature there are tiny flowers found along the spadix but the spathe is not a flower. Aroids are divided into two basic reproductive sections or groups. The first produces a bisexual inflorescence with male and female flowers mixed together along the length of the spadix. The second is known as the unisexual group and includes those aroids that produce an inflorescence with bisexual flower zones. Anthurium species are bisexual.
 Anthurium differ from
Philodendron species since all Anthurium produce perfect flowers
containing both male and female organs while Philodendron produce
imperfect flowers containing only a single sex. When an
Anthurium is "in flower" the reference is to the tiny flowers
containing both male and female sexual parts that grow on the spadix at the
center of the inflorescence. To help prevent self pollination nature has
designed the female flowers to be receptive before the male portion of
the flower
produce their pollen so in most cases an insect must bring pollen from
another plant.
Anthurium differ from
Philodendron species since all Anthurium produce perfect flowers
containing both male and female organs while Philodendron produce
imperfect flowers containing only a single sex. When an
Anthurium is "in flower" the reference is to the tiny flowers
containing both male and female sexual parts that grow on the spadix at the
center of the inflorescence. To help prevent self pollination nature has
designed the female flowers to be receptive before the male portion of
the flower
produce their pollen so in most cases an insect must bring pollen from
another plant.
At female anthesis when the female flowers are produced the spadix produces female receptive flower parts followed by sexually active male and sterile flowers. The flowers are almost microscopic and since they are bisexual grow mixed together in a single flower all along the spadix. If the female flowers are pollinated by an appropriate insect which is normally a beetle from the genus Cyclocephala that carries pollen from another specimen at male anthesis the spadix will begin to grow berries. In many Anthurium species it is possible for the plant to self pollinate and it is theorized the wind may play a part in self pollination.
These pollinators are drawn to the plant since it produces a "perfume" known as an pheromone once it reaches anthesis. The beetles can detect the pheromone from distances as great as 200 meters (600 feet) away and fly a zig-zag pattern through the forest to locate its source since the wind does not follow a straight-line path through the forest. Along with a mate the beetles are drawn by both the fragrance and the heat of both thermogenesis (heat rise) and the "glow" of infrared heat which the beetles can "see" with their antennae in the dark. The beetles are also drawn to the inflorescence since the pollen being produced is a food source in the form of lipid protein.
For more detailed information on how aroid species reproduce please read this link: Aroid pollination
The Inflorescence of Anthurium sagittatum is normally erect but may be re-curved as can be seen in our photos . The spathe is papyraceous (paper-like) and persists (does not quickly fall off) when in fruit. The spathe is ribbed and is green to yellowish green and may be tinged purplish as it ages. The spathe is also matte on the inside and is spreading in shape. The spathe may be tinged reddish purple and is weakly glossy on the outside. The spadix is a medium to pale yellow green and is semi glossy but may change to purplish as it matures.
Joep states that Anthurium sagittatum specimens are often in flower in French Guiana and once pollinated produce deep purple berries (fruit) which contain the seeds of the species. Once pollinated the inflorescence is then known as an Infructescence. The berries are rounded to obovate (oval and to oblong) and are eaten by birds and other rain forest animal which spread the seeds through the rain forest in their droppings.
 In
in several published papers including a study
entitled The Reproductive Biology of Euglossine-pollinated plants in the
natural reserve Nouragues, French Guiana by Heiko Hentrich
done in French Guiana,
the author states
"The reproductive biology of several Euglossine-pollinated species of an
understory community in the natural reserve Nouragues in French Guiana were
studied. The first two chapters deal with the pollination biology of three
sympatric, co-flowering aroids (Anthurium sagittatum, Anthurium thrinax and
Spathiphyllum humboldtii). Each of the studied species possessed a specific
floral scent bouquet that attracted species-specific pollinators. The
pollinators were male Euglossine bees, which collected floral scent at the
spadices of the plants. Floral scent might be a major factor in the reproductive
isolation of these plant species. "
In
in several published papers including a study
entitled The Reproductive Biology of Euglossine-pollinated plants in the
natural reserve Nouragues, French Guiana by Heiko Hentrich
done in French Guiana,
the author states
"The reproductive biology of several Euglossine-pollinated species of an
understory community in the natural reserve Nouragues in French Guiana were
studied. The first two chapters deal with the pollination biology of three
sympatric, co-flowering aroids (Anthurium sagittatum, Anthurium thrinax and
Spathiphyllum humboldtii). Each of the studied species possessed a specific
floral scent bouquet that attracted species-specific pollinators. The
pollinators were male Euglossine bees, which collected floral scent at the
spadices of the plants. Floral scent might be a major factor in the reproductive
isolation of these plant species. "
Within the genera Euglossini there are some 200 Neotropical bee species known as either "Orchid Bees" or Euglossine bees Five genera are known to be found exclusively in Neotropical America (South and Central America). The bees are characterized by a very long tongue and the behavior of the males of a species is known to be drawn to the pheromones (fragrances) from many flowers and other objects such as dead wood. These bees store what they collect on their modified hind legs. Bees in the genera Euglossini include vividly colored bees with colorations of metallic blue, greens and bronzes and the bee species are known to serve as orchid pollinators in South American rain forests. Utilizing their long tongues which measure up to 4 cm (1.6 inches) in length these bees are known to pollinate close to 700 species of tropical orchids. These bee species are are common in lowland forests where the majority of Anthurium sagittatum specimens are found.
However other experts are convinced these bees may only be
drawn to the pheromone of Anthurium species and in fact only infrequently
pollinate aroid species. An observation of any insect visiting a
particular plant species requires careful study before any scientific
conclusions can be drawn.
 The insect may only be drawn to the pheromone
(perfume) and does not actually transfer pollen from one plant at male anthesis to another
at female anthesis. In a personal note from noted aroid
expert Julius Boos he wrote, "Euglossine
bees are small and are about 1/3 the size of a honey bee. These metallic
bees are found in the tropics. They are attracted to special odors given
off by some aroids and orchids, and are assumed to be the pollinators of some
aroids, Spathiphyllum cannifolium (see Deni Bown`s book on this), and maybe some
Anthuriums, etc."
The insect may only be drawn to the pheromone
(perfume) and does not actually transfer pollen from one plant at male anthesis to another
at female anthesis. In a personal note from noted aroid
expert Julius Boos he wrote, "Euglossine
bees are small and are about 1/3 the size of a honey bee. These metallic
bees are found in the tropics. They are attracted to special odors given
off by some aroids and orchids, and are assumed to be the pollinators of some
aroids, Spathiphyllum cannifolium (see Deni Bown`s book on this), and maybe some
Anthuriums, etc."
Although other references can be found indicating Euglossine bees are primary pollinators of Anthurium species it is well known that many Anthurium sp. are capable of self pollination and an insect simply walking along the spadix may transfer pollen from the male flowers to the female flowers. Casual pollination is not the same as a primary pollinator.
To bring some clarity to this subject I asked my friend French aroid pollination expert Dr. Marc Gibernau (GHEE-ber-no) of the University Paul Sabatier in Toulouse, France if Euglossine bees were involved in Anthurium pollination. Marc responded, "Euglossine bees are primary pollinators of many Anthurium but not all." He then continued ".... apparently some species may be beetle pollinated (curculionid weevils) and one species is suspected to be hummingbird pollinated. But pollination ecology in Anthurium is poorly documented". At the end he stated only 15 Anthurium species may be pollinated by bees.
As a result of both Julius' and Marc's notes it may be premature to assume bees from the genera Euglossini are in fact a "primary pollinator" of very many Anthurium species at all. Certainly these bees do visit some Anthurium species but beetles from the genera of Cyclocephala are likely to be primary in the pollination process of the majority of Anthurium species. The jury is still out on this subject.
Julius'
mention of Deni Bown's
book is a reference to her excellent text Aroids, Plants of the Arum Family
which is available at many book stores.
Our specimen shown in all the photos on this page was acquired from the
collection of the Missouri Botanical Garden in St. Louis, MO. It was
grown from an offset of specimen 81084 which was originally tagged
Anthurium rubrinervium.
The
specimen was collected by Dr. Tom Croat and was
presented to us on June 19, 2009.
For a better understanding of variation within plant species please click this link Natural variation within Aroid and other plant species.
Looking for a specimen? Contact
http:///
